Carro
Carro

Serotonin
26 Dec 2016
Serotonin è l'ammina biogenic che è formata di amminoacido tryptophan dal suo hydroxylation e decarboxylation. Una quantità importante di un serotonin contiene nelle celle enterokhromaffinnykh d'intestino, CNS, principalmente in hypothalamus e mesencephalon, thrombocytes, più piccola quantità nel labrotsitakh, le celle di albero, le surrenali. Un serotonin esercita l'effetto su attività nervosa, riduzione di cause di musculation liscio di un intestino, un utero, bronchi, e anche restringersi di navi. Le reazioni di un organismo a un serotonin sono la pietra angolare il centrale, myotropic, ganglionic, gli effetti riflessi.
Istruzione. Un serotonin (5-gidroksi-triptamin, 5-HT) è sintetizzato nelle celle di epitelio intestine enterokhromaffinnykh da un L-tryptophan. Un serotonin è formato anche in celle nervose di mezenterialny plexus e in un CNS dove i giochi un ruolo di mediatore. Thrombocytes non sintetizzano un serotonin, comunque lo prendono e si accumulano.
Recettori di Serotonin. Ci sono parecchi tipi di recettori. Dai recettori di punto di vista farmacologici 5-HT1 e 5-NT2, e anche 5-HT3 e 5-HT4 sono importanti. Di regola, i recettori lavorano attraverso G-proteina. Il recettore 5-HT3 rappresenta il canale cationic.
Effetto di serotonin. Sistema cardiovascolare. L'influenza di serotonin su sistema cardiovascolare è complessa siccome il serotonin ha l'effetto (qualche volta di fronte mandato) in molti siti attraverso tipi diversi di recettori. Attraverso 5-HT2-retseptory situato in muscoli instriati di pareti di navi, un serotonin causa restringersi di navi. L'effetto nello stesso momento indiretto di serotonin può condurre a vasodilatation e depressione dell'ABP: attraverso 5-HT1A-retseptory blocca neuroni compassionevoli di un brainstem e la periferia che conduce a depressione di tonus compassionevole; l'attivazione di recettori 5-HT1 in endothelium vascolare conduce a emissione di vazodilatator (prostatsickin, NESSUNO). Il serotonin che è rilasciato da thrombocytes gioca un ruolo in processi di thrombogenesis, hemostasis e in hypertonia pathogenesis a donne incinte.
Sumatriptan (un agente contro emicrania) è un 5-HT1D agonist e 5-HT1B-retseptorov. Restringersi di navi craniche causate da blocco di emissione di neuropeptids conduce a un cosiddetto �neurogenic l'infiammazione� o dirigere la crisi vascolare. Nello stesso momento ci può essere un sentimento di costrizione in un petto legato a una critica spastica di arterie coronarie. Altri �tryptones� sono naratriptan, zolmitriptan e rizatripan.
Distesa digestiva. Il serotonin formato in mezenterialny plexus o celle enterokhromaffinnykh stimola un peristalsis e una secrezione di un intestino per mezzo d'influenza su 5-HT4-retseptory.
I tentativi di effetto farmacologico su un motility di un intestino attraverso recettori serotonin sono stati inefficaci finora. Tsizaprid, un 5-HT4 agonist, comunque, può aumentare motility propulsivo di un intestino. Questo effetto collaterale è bloccato da Atropinum che dimostra l'aumento di concentrazione di Acetylcholinum sotto l'influenza di tsizaprid. Il farmaco rende una serie di effetti collaterali. È inactivated sul meccanismo con partecipazione di SURZA4 che conduce a varie interazioni impreviste con altre sostanze. Tsizaprid causa disordini (compreso serio) un ritmo cordiale (la riduzione di intervalli QT) perciò il farmaco non ha trovato l'applicazione.
Sistema nervoso centrale. I recettori di Serotonin giocano un ruolo importante nelle varie funzioni CNS.
Fluoxetine è l'antidepressivo che blocca il ritorno serotonin la ricomprensione. Considerevolmente aumenta la motivazione, e anche abbassa l'appetito.
Sibutramin, blocker del ritorno neuronal serotonin ricomprensione e Noradrenalinum; è applicato come rimedio per obesità.
Ondansetron ha l'effetto di antiemetico espresso al vomito provocato da applicazione di tsitostatik. È un antagonista di 5-HT3-retseptorov. Gli analoghi di ondansetron sono tropisetron e granisetron.
L'acido e gli altri di psychedelia (psikhotomimetika), come mescalina e psilocybin, provocano allucinazioni, i disordini di coscienza, paura, forse, a causa di attivazione di 5-NT-retseptorov.
Effetti di serotonin
Il personale di università di McMaster è diventato autori di lavoro scientifico durante cui il gene chiave di obesità - una sorta di serotonin è stato trovato. È ben noto che serotonin è �l'ormone di gioia�, il suo sviluppo da un cervello promuove la stabilità emotiva e il buon umore. Ma, come i ricercatori canadesi spiegano, il serotonin che è responsabile di emozioni piacevoli appartiene al primo tipo di questa connessione.
�È diviso in due tipi: nella scena di azione e in una forma di sintesi. Il primo tipo è sviluppato in un cervello e influenza emozioni diverse�, - i biologi hanno spiegato.
Al secondo tipo, serotonin periferico appartiene questa sostanza regola l'attività di tessuto adiposo marrone da cui lo sviluppo di obesità dipende. Può anche piacerLe Pankramin.
Il grasso marrone contiene componenti che promuovono la diminuzione in zucchero in sangue, e anche a bruciato di calorie e la loro elaborazione in energia. Su un corpo della persona ci sono zone certe dove il grasso marrone si calma e le sue gabbie sono più attive, la cifra alla persona è più armoniosa. I ricercatori canadesi sono venuti a conclusione che sopprimendo serotonin del secondo tipo; è possibile aumentare l'attività considerevolmente metabolica di celle di tessuto adiposo marrone. A sua volta, costringerà un corpo �a bruciarsi� il grasso bianco � e, succederà indipendentemente da come il cibo di caloria alta è mangiato dalla persona.
Recettori di Serotonin
Gli effetti di serotonin sono estremamente vari. Questa sostanza i servizi come un mediatore in TsNS, influenza muscoli lisci di navi e un IDIOTA, partecipa a trombotsitarnom vascolare un homeostasis. I metodi di clonazione molecolare hanno rivelato un improvvisamente gran numero i recettori serotonin che si sono basati su struttura e la funzione può esser divisa in 4 tipi. 5-T1--, 5-HT2-and 5-HT4-retseptory è integrato a G-proteine e attraverso queste proteine e i sistemi corrispondenti dei secondi mediatori influenzano funzioni di vari enzimi e le proprietà elek-trofiziologich di gabbie effector. Anzi, 5-HT3-retseptory sono legati a canali d'ione. Qui considereremo stimulators e blockers i recettori serotonin. Le più nuove medicine di questi gruppi che in modo selettivo fanno funzionare su subtipi separati i recettori serotonin sono state ricevute in lavori con uso di recettori recombinant. Ci fermeremo anche su modelli sperimentali che si rivolgono a una ricerca dei mezzi che influenzano funzioni mentali difficili e la loro persistenza di violazioni, comportamento aggressivo, disagio, depressione, insonnia di sogno di ciclo e altro. stimulators selettivi moderni di subtipi separati i recettori serotonin già con successo sono applicati in caso di emicrania e disagio e blockers selettivo in caso di parecchie violazioni gastrointestinali. È possibile influenzare effetti fisiologici di serotonin anche per mezzo dei mezzi che operano a trasferimento di serotoninergich. Dunque, gli inibitori della cattura di ritorno di serotonin sono stati medicine efficaci per cura di depressione e disagio.
Nonostante il fatto che il ruolo serotonin in molti processi fisiologici e patologici non sollevi dubbi, i punti della sua applicazione e i meccanismi di azione sono studiati male. Forse, una tale situazione è parzialmente causata da varietà il serotoninovykh di recettori. Questi recettori registrati all'inizio da metodi farmacologici sono ricevuti da KDNK che clona oggi. I recettori di Recombinant serotoninovy usano per studiare di meccanismi molecolari di effetto di serotonin, e anche per ricerca degli agenti che in modo selettivo influenzano subtipi separati di questi recettori. Il cerchio di un uso clinico di agenti simili diventa più largo e più largo.
Informazioni storiche
In 1930esimo Erspamer ha cominciato a studiare la localizzazione di celle enterokhromaffin per mezzo di macchie su derivati indole. La concentrazione più alta di tali derivati è stata registrata in mucoso da un IDIOTA; più lontano ci sono stati thrombocytes e alcuni reparti di un CNS (Erspamer, 1966). Poco dopo Peydzh e coautori, che lavorano nella clinica di Clevelend, per la prima volta emisero la sostanza vasoconstrictive rilasciata da thrombocytes nel corso di un arresto di sanguinamento e decifrarono la sua struttura (Il rapporto e al., 1948). Questa sostanza chiamata da Paige un serotonin (Page, 1976), ha apparito a quei derivati di un indole che è stato investigato da Erspamer. La descrizione di strade di sintesi e disintegrazione di un serotonin (l'Uden-amico, 1959) e le sue proprietà angiotonic (Sjoerdsma, 1959) ha permesso di fare un'ipotesi secondo cui le implicazioni di una sindrome carcinoid cosiddetta a pazienti con tumori da celle enterokhromaffin sono causate dalla produzione aumentata di questa sostanza. Veramente, tali pazienti hanno un'espulsione quotidiana con urina di un serotonin e il suo metabolites può arrivare a centinaia di milligrammi. Alcuni sintomi di questa malattia fino a un certo grado indicano meccanismi di effetto di serotonin. Dunque, a pazienti le psicosi simili all'acido che si alza a ricevimento possono svilupparsi. Pensando che in animale e sostanze di tessuti vegetali con azione allucinogena sono trovati simili con tripta-miny, è possibile presumere che le sostanze simili sono formate e causano una sintomatologia psicotica in pazienti con sindrome characinoid. Su funzione di mediatore di un serotonin in un cervello di mammiferi è stato suggerito a metà degli anni cinquanta (Brodie e Shore, 1957).
I primi dati su meccanismi molecolari di effetto di serotonin sono stati ottenuti in esperienze in un fegato di Fasciola hepatica (Mansour, 1979). Sotto l'influenza di serotonin a esso la mobilità e la concentrazione di tsAMF bruscamente sono aumentate; sia questo, sia l'altro effettua sono stati bloccati da un acido. L'aumento di mobilità è stato causato da phosphorylation tsAMF-dipendente di fosfofruktokinaza che limita glycolysis l'enzima. Comunque i recettori serotoninovy che mediano questi effetti a un fegato, probabilmente, altri, che recettori di mammiferi connessi con adenilattsiklaz a serotonin. Al più tardi così i dati dettagliati su meccanismi di effetto di serotonin non è stato successo per ricevere finora.
Serotonin ha apparito su impianti e animali già in prime fasi di evoluzione, e a questi, forse, abbondanza di recettori serotonin parla (Peroutka e Howell, 1994). La clonazione di questi recettori ha mostrato che alcune medicine che sono state ancora prima considerate selettive nei loro subtipi separati veramente hanno l'affinità alta in relazione a parecchi subtipi. In modo più dettagliato su storia di studio ed effetti di serotonin vedi l'articolo Sjoerdsma e Palfreyman (1990).
Proprietà chimiche di serotonin
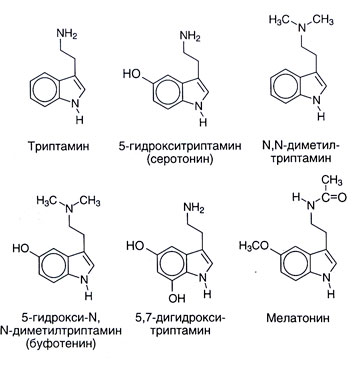
Fonti. La struttura chimica di serotonin e alcune connessioni vicine con lui è data in fico. 11.1. Serotonin è molto diffuso in vita di animale e un impianto: è trovato a vertebrato, tunicates, molluschi, arthropods, coelenterates, in frutto e in noci. È trovato anche in veleni � in un'ortica, in vespe e scorpioni. I parenti sintetici o naturali numerosi a sostanza serotonin anche in un vario grado hanno gli effetti fisiologici centrali e periferici. Molti Né O-metilirovannye indolamines (per esempio, N, N-dimetiltriptamin) sono hallucinogens. Siccome possono esser sviluppati in un organismo, a lungo li ha considerati possibile responsabile di almeno alcune esposizioni di psicosi. La melatonina (5 metoksi N atsetiltriptamin) è formata di serotonin da N-acetylation con O-methylation successivo (il fico. 11.2). Questa sostanza serve come indolamine principale di corpo dove la sua sintesi è regolata da fattori esterni (soprattutto, il livello d'illuminazione). La melatonina causa depigmentation di melanotsit di pelle e sopprime la funzione di ovaie. Forse, recita una parte in bioritmi e perciò può essere utile a una sindrome di cambiamento di fusi orari.
Sintesi e catabolismo. Un serotonin è formato di amminoacido insostituibile di un tryptophan in 2 stadi (il fico. 11.2). Al primo stadio sotto l'influenza di triptofangidroksilaza 5 hydroxytryptophan sono formati, è la limitazione serotonin la reazione di sintesi. Triptofangidroksilaza rappresenta un oxidase con le funzioni di admixed. L'ossigeno molecolare, e come coenzyme tetrahydrobiopterine prende parte alla reazione catalizzata da esso. L'attività di triptofangidroksilaza, così come tyrosinehydroxylase, è regolata da phosphorylation, comunque il triptofangidroksilaza non è interdetto dal prodotto finito sul meccanismo di feedback negativo. In un cervello di triptofangidroksilaz non è saturato con substrato e perciò il tasso di sintesi di un serotonin dipende da concentrazione di un tryptophan. L'ultimo viene a celle cerebrali da cattura attiva per mezzo del corriere che è responsabile di trasporto di parecchi amminoacidi neutrali e ramificati. A questo proposito la manutenzione di un tryptophan in un cervello dipende non solo dalla sua concentrazione in plasma, ma anche su concentrazione di altri amminoacidi che competono con un tryptophan per un corriere.
Decarboxylation conduce 5 hydroxytryptophans a formazione di un serotonin. La disputa lunga su se decarboxylases 5 hydroxytryptophans e DOFA sono diversi o lo stesso enzima, è stata risolta da KDNK i metodi clonanti si è scoperto che lo stesso prodotto di gene è responsabile di decarboxylation di entrambi i substrati. Adesso questo enzima è chiamato decarboxylase di L-amminoacidi aromatici. È estremamente molto diffuso e intacca molti substrati. 5 hydroxytryptophan sono decarboxylized molto rapidamente e in un cervello non è quasi trovato. A questo proposito i tentativi di intaccare la concentrazione in un cervello serotonin da concentrazione cambiano 5 hydroxytryptophans sono destinati a fallimento.
La strada principale di catabolismo serotonin la trasformazione in acido 5-hydroxyindolacetic che anche procede in 2 stadi (il fico. 11.2). All'inizio sotto l'influenza di MAO il 5-hydro-xyindolacetaldehyde che allora passa in acido 5-gidroksi-indolacetic sotto l'influenza di enzima aldegiddegidrogenaza molto diffuso in un organismo è formato (la quantità insignificante che 5 hydroxyindolacetaldehydes trasformano in alcool � 5-gidroksitriptofol). L'acido 5-hydroxyindolacetic è activly tolto da un cervello; questo processo è soppresso con un inibitore nonspecifico di probenetsidy di trasferimento di transepithelial. Come in celle nervose il quasi 100% di tutto metabolites di un serotonin cade alla parte di acido 5-hydroxyindolacetic, il tasso di un giro di un serotonin in un cervello è valutato ad aumento di livello di acido 5-hydroxyindolacetic dopo introduzione di probenetsid. L'acido 5-hydroxyindolacetic che è formato in un cervello e altri organi, e anche piccole quantità di un 5-gidroksitriptofol e glucuronides è tolto con urina. Normale il quotidiano egestion di acido 5-hydroxyindolacetic all'adulto fa 2 � 10 mg. Valori più alti � un segno affidabile di sindrome carcinoid. La sintesi bruscamente aumentata di un serotonin a questa malattia esige un gran numero di pyridinic nucleotides e un tryptophan e perciò segni d'insufficienza di acido nicotinic e un tryptophan non una rarità a tali pazienti. L'aumento di cause di etanolo di manutenzione di NADN, e come risultato di 5 hydroxyindolacetaldehyde passa da una strada ossidante di un catabolismo a ricupero (il fico. 11.2).
Questi sono aumenti egestion di un 5-gidroksitriptofol un po' e rispettivamente riduce egestion di acido 5-hydroxyindolacetic.
Ci sono due isoenzymes di MAO: MAO A e MAO B. All'inizio sono stati divisi basati su affinità a sostrati e sensibilità a inibitori; adesso sia gli isoenzymes sono clonati, sia le proprietà delle forme clonate e naturali sono state identiche (Shih, 1991; vedi anche hl. 10). MAO A ha l'affinità preferenziale a serotonin e noradrenaline, e il suo inibitore selettivo è clorgilin. MAO B più lavori su β-fenil-etilamin e benzylamine; MAO B inibitore selettivo selegilin. L'affinità sia di isoenzymes a dopamine sia di triptamin è identica. Le gabbie nervose contengono sia MAO A sia MAO B � principalmente su una membrana esterna di mitochondrions. Siccome isoenzyme principale di piastrine che anche contengono serotonin in concentrazione alta serve a MAO B.
Si ha supposto che ci siano anche altre strade di un catabolismo di serotonin, per esempio sulphation e Su - o N-methylation. Il viaggio finale, soprattutto, poteva condurre a formazione di una sostanza psychotropic endogena 5-hydroxy-N, N-dimetiltriptamina (bufotenina, il fico. 11.1). Comunque altri metilirovanny indolamines (N, N-dimetiltriptamin, 5-metoksi-N, N-dimetiltriptamin) hanno la gran parte più espresso proprietà allucinogene, e il loro ruolo in pathogenesis di psicosi è più probabile.
L'inactivation di serotonin è compiuto non solo da disintegrazione enzymatic, ma anche per mezzo della cattura di ritorno. Na + - il corriere dipendente situato su una superficie esterna di membrana presynaptic della fine serotoninergich (provvede l'eliminazione di serotonin da un'incrinatura sinottica) e una superficie esterna di una membrana di piastrine è responsabile di questa cattura (estrae serotonin da sangue). A piastrine è la strada unica di azione di rifornimento di serotonin siccome non ci sono enzimi di sintesi di questa sostanza in loro. Il corriere serotonin, così come altri corrieri di monoammine, è clonato (hl. 12).
Punti di applicazione serotonin
La maggior parte parte di tutta la manutenzione di serotonin in un organismo cade alla parte di tessuti peripheric sebbene serva anche come un mediatore in un CNS. La sua concentrazione in celle enterokhromaffinnykh e thrombocytes è la più alta. Un serotonin gioca un ruolo importante in una regolazione di un motility di un IDIOTA.
Celle di Enterokhromaffinny. Queste celle calmano in mucoso un IDIOTA. Particolarmente ci sono molti di loro in un duodeno. Nelle celle enterokhromaffinnykh è sintetizzato da un tryptophan e un serotonin si riunisce, e anche altri agenti biologicamente attivi, per esempio la sostanza P e kinina contengono. C'è un livello certo di secrezione basale di un serotonin in un IDIOTA. Questa secrezione amplifica all'atto di stiramento meccanico (per esempio, entrando in una nutrizione o una soluzione salina ipertonica) e a una noiosa di fibre di motivo di nervi vagus. Forse, l'effetto stimolante di serotonin su un motility di un IDIOTA è mediato anche dalla sua influenza su neuroni di plexus intermuscolare (Gershon, 1991; vedi anche hl. 38). La secrezione bruscamente aumentata di un serotonin e altri agenti biologicamente attivi a sindrome carcinoid è seguita dai disturbi gastrointestinali, cardiovascolari e nervosi corrispondenti. Inoltre, la sintesi aumentata di un serotonin può avere come conseguenza l'insufficienza di acido nicotinic e un tryptophan.
Thrombocytes. Thrombocytes differiscono da altri elementi formulati di un sangue, soprattutto, in capacità di prendere il deposito e rilasciare un serotonin. La sintesi di un serotonin in thrombocytes non avviene. Un serotonin è preso thrombocytes da un sangue e viene per immagazzinamento a segreto elektronop granelli per mezzo di trasporto attivo. Questi processi sono in molti aspetti simili a cattura e immagazzinamento di Noradrenalinum nelle fini compassionevoli (hl. 6 e 12). Attraverso una membrana di thrombocytes un serotonin è trasferito per mezzo di Na + - il trasporto dipendente, e a granelli � da trasporto attivo secondario con uso come una fonte di energia di una pendenza elettrochimica per H + incorniciato da N +-Atfazoy. Nello stesso momento la concentrazione di un serotonin in granelli arriva a 0.6 mol/l, è 1000 volte più alto, che in citoplasma di thrombocytes. Tasso Su + - ricomprensione serotonin dipendente da thrombocytes un indicatore sensibile di attività di inibitori di ricomprensione serotonin.
La funzione principale di thrombocytes è hemostasis: chiudono spazi vuoti in endothelium danneggiato. D'altra parte, l'integrità di un endothelium gioca un ruolo importante in funzionamento di thrombocytes (Furchgott e Vanhoutte, 1989). L'endothelium costantemente si mette in contatto a thrombocytes come a causa delle forze di cambiamento che agiscono nel sangue attuale sono spostati alla periferia di navi (Gibbons e Dzau, 1994). L'effetto di Vasoconstrictive di serotonin e tromboksan di A2 sono resistiti da un fattore endoteliale di vasorelaxation (N0 e, forse, altre sostanze) (Furchgott e Vanhoutte, 1989; fico. 11.4). Per adesione e aggregazione di thrombocytes la condizione di un endothelium ha l'importanza cruciale (Hawiger, 1992; Ware e Heistad, L993). Quando thrombocytes confinano a endothelium danneggiato, emettono le sostanze che causano la loro adesione e rilascio di un serotonin. La RADIOGONIOMETRIA AUTOMATICA e ò ð îìáîêñàí A2 appartengono a tali sostanze (hl. 26 e 55). Linkng di un serotonin con 5-HT2A-retseptorami ha l'effetto proagregant debole che bruscamente amplifica alla presenza di un collagen. Se il difetto di una parete vascolare arriva a strati muscolari lisci, allora un serotonin rende l'effetto vasoconstrictive diretto che serve come uno di meccanismi hemostasis. Questo effetto amplifica sotto l'influenza di agenti biologicamente attivi che sono emessi nel campo di danno tromboksan di A2, kinin, vasoactive peptides. La formazione di thrombi a un atherosclerosis è promossa da distruzione di un endothelium e, come risultato, la mancanza di un fattore endoteliale di vasorelaxation. In queste condizioni la conduzione di processi a thrombogenesis procedono incontrollatamente, come un circolo vizioso. Una parte in loro è recitata anche da un serotonin. Il quadro simile può esser osservato ad altre malattie di navi, per esempio la sindrome di Reynaud e vasospastic stenocardia.
La descrizione a fico. 11.4. Funzioni di un serotonin di thrombocytes. Il rilascio di un serotonin da thrombocytes è iniziato loro da adesione e aggregazione. A sua volta, un serotonin causa 1) l'attivazione á-ÍÒ-receptors thrombocytes e, vrezultata, il cambiamento di una forma e l'accelerazione di aggregazione dell'ultimo, 2) l'attivazione 5-HT, - i recettori simili di un endothelium con allocazione di un fattore endoteliale di vasorelaxation, 3) l'attivazione di S-HT-receptors di muscoli instriati di navi e restringersi dell'ultimo. Tutti questi processi procedono in interazione in molti altri agenti biologicamente attivi e alla fine conducono a sanguinamento di arresto.
Sistema cardiovascolare. La reazione tipica di vasi sanguigni a serotonin si restringe. Le navi di organi d'IDIOTA, reni, polmoni e cervello sono particolarmente sensibili a esso. Serotonin causa anche la riduzione di muscoli instriati di bronchi. I suoi effetti su cuore sono vari che il cambiamento di recettori serotonin di tonus di nervi vegetativi e reazioni riflesse è spiegato da attivazione di subtipi diversi (Saxena e Villalon, 1990). Dunque, chronotropic positivo diretto ed effetto inotropic di serotonin su cuore possono esser mascherati da effetti di esaltazione delle fibre che vanno da recettori di bar e chemoceptors. L'influenza di un serotonin alle fini afferent di nervi vagus causa Yarisha riflesso di Betsold che è mostrato bradycardia affilato e caduta dell'ABP. Qualche volta gli arterioles sotto l'influenza di un serotonin non sono restretti, e, anzi, la soppressione di rilascio di Noradrenalinum dalle fini compassionevoli si estendono come risultato di allocazione di un fattore endoteliale di vasorelaxation e Prostaglandinums, e anche. D'altra parte, un serotonin in sé rafforza l'azione vasoconstrictive di Noradrenalinum, angiotensin 11 e Histaminum. Promuove l'effetto statico haemo ancora più efficace di serotonin (Gershon, 1991).
IDIOTA. Probabilmente, siccome la fonte principale e l'immagazzinamento di un serotonin in un organismo servono celle enterokhromaffinny mucose un IDIOTA. Il serotonin assegnato da queste celle supera una vena portale a un fegato dov'è metabolized sotto l'influenza di MAO A (Gillis, 1985). Alcuna quantità di serotonin passa un metabolismo epatico, ma è rapidamente presa un endothelium di vasi capillari polmonari e anche è intaccata da MAO. Il serotonin che è assegnato in una parete di organi di un IDIOTA all'atto del loro stiramento meccanico o esaltazione di nervi vagus partecipa a una regolazione locale di questi organi. Sotto l'influenza di un serotonin un motility di uno stomaco e un intestino può sia amplificare, ed esser frenato (Dhasmana e al., 1993) come in un IDIOTA ci sono almeno 6 subtipi i recettori serotonin (l'etichetta. 11.2). L'effetto stimolante di serotonin è causato dalla sua azione alle fini dei nervi adatti a strati muscolari longitudinali e circolari (5-HT4-retseptory), su neuroni interni (5-HTj-and 5-NT|R-retseptory) e è diretto su muscoli lisci (5-ÍÒ-receptors in intestini e 5-HT2B-retseptory in giorno di uno stomaco). In un esofago il serotonin intacca 5-HT4-retseptory che a tipi diversi di animali può esser seguito sia calando, sia il rilassamento di muscoli lisci. I 5-HT3-retseptory (sono molti visitatori alle fini di fibre sensibili del vagabondaggio e altri nervi, e anche sulle gabbie) giocano un ruolo chiave in un riflesso di emetico (Grunberg e Hesketh, 1993). In una struttura intermuscolare le fini serotoninergetic sono trovate. Il rilascio di serotonin in intestini è causato da acetilcolina, irritazione di nervi compassionevoli e aumento di pressione intestinale intra e diminuzione in pH (Gershon, 1991). Il serotonin che è emesso nello stesso momento, a sua volta, inizia la riduzione di peristaltic.
L'area principale di concentrazione di corpi di neuroni serotonin in CNS è nuclei di cucitura di brainstem. I processi di questi neuroni vanno a tutti i reparti di un midollo spinale e principale (hl. 12). Un serotonin è assegnato non solo nelle fini presynaptic, ma anche in varicosity cosiddetto di axons dove non ci sono sinapsi esattamente espresse (Descarries e al., 1990). In questi casi intacca molte strutture adiacenti subito. Una tale caratteristica di allocazione e un effetto di serotonin saranno composti con il punto di vista molto diffuso che un serotonin non è solo un mediatore, ma anche il modulatore di neurone (hl. 12).
Nelle fini di neuroni serotonin ci sono tutti i componenti necessari per sintesi di un serotonin da un tryptophan (il fico. 11.2). serotonin formato rapidamente viene a bolle sinottiche dove su lui MAO non può lavorare. Dopo rilascio in una fenditura sinottica un serotonin è di nuovo preso la fine nervosa per mezzo di Na - un corriere dipendente. Questa cattura di ritorno serve come una strada efficace di un inactivation di un mediatore. Le stesse molecole di un serotonin che non ritornano alla fine nervosa sono fatte esplodere da MAO situato in neuroni postsynaptic e le prossime celle.
Effetti di Electrophysiological. Questi effetti di un serotonin differiscono in aree diverse di un cervello e in neuroni diversi e dipendono sui quali recettori intacca (l'etichetta. 11.3; Aghajanian, 1995). Un serotonin può avere l'eccitazione e frenare l'effetto che differisce su dinamica provvisoria sugli stessi neuroni. Dunque, in neuroni hippocampus un serotonin causa all'inizio la polarizzazione hyper (causato da attivazione di 1A-recettori 5-HT), allora la depolarizzazione lenta (causato da attivazione di 5-HT4-retseptorov).
TsNS. Serotonin influenza TsNS molte funzioni, compreso un sogno, un'attività cognitiva, una percezione, una direzione di movimenti, controllo termico, sensibilità dolorosa, appetito, comportamento sessuale e regolazione endocrina. In un cervello tutti hanno clonato i recettori serotonin sono trovati, e spesso allo stesso reparto ci sono diversi recettori. Per di più, sebbene l'espressione di recettori serotonin in neuroni separati sia studiata insufficientemente, è possibile credere che sullo stesso neurone parecchi subtipi di questi recettori possono esser situati, e la loro attivazione può esser seguita sia da synergistic sia da effetti antagonistici. Può essere la causa di una varietà straordinaria di influenze di serotonin su funzioni cerebrali.
La polarizzazione di Hyper e la diminuzione in resistenza di una membrana che si alza in caso di attivazione 5-HT) gli A-recettori, sono causati da aumento di permeabilità di potassio. Questi effetti sono bloccati da tossina, ma non dipendono da tsAMF. Perciò, possono esser causati da interfaccia diretta di 5-NT|A-retseptorov (attraverso la G-proteina simile a Gi) con il canale di potassio (Andrade e al., 1986). L'attivazione dei recettori situati su un corpo e dendriti di neuroni di gherigli di una cucitura di un tronco di un cervello anche conduce a K + - la polarizzazione hyper dipendente. Qui anche partecipa a una trasmissione j di un segnale da un recettore al canale sensibile a tossina di G-proteina, ma la corrente di potassio ha altre caratteristiche, che quello che si alza in caso di attivazione di postsynaptic 5-HT1A-retseptorov in un hippocampus. Non si sa già per mezzo quale attivazione di meccanismi 5-HT1D-ayTopeuerrropoB conduce a soppressione di rilascio di serotonin. Probabilmente, il numero dei canali calcic che si aprono in risposta a impulso nervoso le diminuzioni potenziali.
L'attivazione di 5-HT2A-retseptorov è seguita da depolarizzazione lenta. In alcuni reparti di un cervello (per esempio, in corteccia prefrontale, un gheriglio adiacente e un gheriglio di motivo di un nervo facciale) questa depolarizzazione è causata da diminuzione in permeabilità di potassio (Aghajanian e al., 1987). C'è anche altro meccanismo collegato a influenza su canali d'ione; conduce a crescita di eccitabilità di neurone e rinforzo di reazione a mediatori emozionanti (per esempio, un glutamate). Il ruolo di sistema fosfoinozitid in questi effettua non è istituito già. Quando sullo stesso neurone ci sono anche 5-HT1-retseptory e 5-Nt2d-retseptory, la reazione finale a serotonin dipende da un rapporto tra la polarizzazione hyper causata da attivazione di 5-HT1-retseptorov e la depolarizzazione causata da attivazione di 5-HT2A-retseptorov. Sullo sfondo di polarizzazione 5-Nt2d-blokatorov hyper amplifica. In molte aree di corteccia 5-HT2A-retseptory sono situati su neuroni inseriti di Gamkergichesky e su neuroni piramidali. Perciò, la stimolazione di questi recettori può condurre a influenze multidirezionali su neuroni piramidali in dipendenza perché se l'azione su questi neuroni o su neuroni di Gamkergichesky prevarrà. L'attivazione di 5-Nt2s-retseptorov su oocytes di rana, l'espresso di m di RNA di questi recettori, conduce a soppressione di corrente di potassio. In un cervello un tal effetto non è rivelato già. La stimolazione di 5-HT4-retseptorov che è seguito da attivazione anche causa la depolarizzazione lenta causata da diminuzione in permeabilità di potassio in neuroni. Non è chiaro già perché due tipi diversi di recettori serotonin connessi a vari sistemi dei secondi mediatori rendono lo stesso effetto fisiologico. Per di più, la depolarizzazione lenta è causata da attivazione di ancora uno tipo di recettori serotonin 5 HT1P-recettori. Questi recettori sono accompagnati da disponibile solo su neuroni intestinali intra e hanno proprietà farmacologiche speciali (Gershon, 1991).
L'attivazione di 5-HT3-retseptorov causa la depolarizzazione bystry. È causato aprendosi del canale che passa Na + e K + (Higashi e Nishi, 1982) e fa un complesso uniforme (hemo il canale sensibile) con un recettore. L'organizzazione simile di un simile 5-HT3-retseptora all'organizzazione N-holinoretseptora è stata confermata in esperienze con fissaggio locale. I 5-HT3-retseptory sono trovati in TsNS, gangli compassionevoli, fibre afferent paracompassionevoli e compassionevoli, intra i neuroni intestinali e le linee cellulari di origine neyronal (per esempio, NG108-15). Sulle proprietà farmacologiche 5-HT3-retseptory differiscono da altri serotonin recettori; forse, ci sono parecchi subtipi di questi recettori che differiscono in varie combinazioni di subjunits.
Funzioni mentali. I mezzi che operano a recettori serotonin e vari cambiamenti di funzioni mentali. Molti modelli sperimentali intesi per una stima provvisoria dell'incentivazione o il bloccaggio di attività di medicine in relazione a questi o quei recettori, sono basati su una ricerca di tali atti di motivo stereotypic come, per esempio, il riflesso. Le tecniche di Behaviouristic (per esempio, un metodo di scelta di medicina) permettono di suggerire su un componente soggettivo di reazioni a farmaci psychotropic. Queste tecniche sono usate anche per una ricerca delle medicine che operano a trasferimento di serotonin, in hallucinogens particolare (vedi sotto). L'analisi di numero enorme dei lavori dedicati a influenza di serotonin su comportamento è al di là del nostro libro, e ci fermeremo solo su quegli esperimenti che hanno un portamento diretto su psychopathology della persona. Per conoscenza più dettagliata con questo soggetto è possibile raccomandare alle rassegne eccellenti Glennon e Lucki (1988), Zifa e Fillion (1992), Koeketal. (1992).
Vada in bicicletta un sogno è l'insonnia. La regolazione di un ciclo un sogno è l'insonnia è diventata una delle prime funzioni mentali per cui il ruolo serotonin è stato esattamente istituito. Dopo il lavoro classico su gatti che è effettuato da Mouret e ñîò ð. (Mouret e al., 1967), la serie di dati che l'esaurimento di provvista serotonin per mezzo di cause l'insonnia eliminata entrando di un predecessore di serotonin 5 di appariti. Ha sembrato anche che tryptophan e non stimulators selettivo di recettori serotonin accorciano il tempo di addormentarsi ed estendono la durata generale di un sogno. Blockers di recettori serotonin può sia alzarsi, e abbassarsi una parte di un sonno lento profondo che, probabilmente, è causato da azione su subtipi diversi di recettori serotonin (Wasquier e Dugovic, 1990). Sia ad animali sia a blockers 5-HT2A-and 5-Nt2s-retseptorov (per esempio, riganserin) piuttosto in modo affidabile causano l'aumento di una parte di un sonno lento profondo.
Aggressione e impulsività. I dati hanno esistito sia su animali sia sulla persona, dimostri che un serotonin gioca un ruolo importante in comportamento aggressivo e impulsivo. In molta comunicazione di prove cliniche tra il livello basso di 5 - di acido hydroxyindolacetic in SMZh e un tal comportamento è mostrato (Brown e Linnoila, 1990). Dunque, la depressione di questo livello è legata a tentativi impulsivi di suicidio (ma non con pensieri suicidi; Virkkunen e al., 1995). Così come in caso di tutti gli altri effetti di un serotonin, gli animali non hanno dati finali su influenza di un serotonin su comportamento aggressivo già sebbene la supposizione di una tal influenza sia abbastanza provata. Recentemente ci sono stati sostegno di dati genetico e dilatazione di tali viste. I recettori di Serotonin investigati da metodi d'ingegneria genetica 5-HT) il V-recettore è stato il primo di. Da una ricombinazione omologa la linea di topi che hanno un gene che cifra questo recettore è stata ricevuta è stato inactivated (Saudau e al., 1994). A tali animali l'aggressione più affilata si è sviluppata quello che parla su un ruolo di questi recettori o in accadere dei contorni di neuronic che sono responsabili di comportamento aggressivo o è immediato in la maggior parte tal comportamento. Alla persona la mutazione punteggiata del gene che cifra MAO A è registrata; l'aggressione nello stesso momento estrema in combinazione con ritardo mentale è anche osservata (Brunner etal., 1993). Come ha apparito, le implicazioni corrispondenti sono disponibili anche per topi con le mutazioni che hanno come conseguenza l'insufficienza di MAO A (I casi e al., 1995). Questi dati, certamente, confermano un'ipotesi di un ruolo di disordini di trasferimento di serotoni-nergic in comportamento aggressivo.
Disagio e depressione. Rendimento dei mezzi che influenzano serotoninergichesky trasferimento (per esempio, inibitori della cattura di ritorno di serotonin), in caso di frustrazione inquietante e depressione la prova per vantaggio di un ruolo serotonin in pathogenesis di queste condizioni. Comunque su modelli sperimentali classici di questi la frustrazione riceve risultati ambigui l'effetto di medicina dipende da specie e razza di un animale e la tecnica applicata. Dunque, agonist parziale di 5-HT1A-retseptorov buspiron (hl. 19), essendo un tranquillante efficace alla persona, non cala a disagio di animali nelle esperienze date da una tecnica di preferenza o evitare; nel frattempo questa tecnica è stata usata in caso di sviluppo di tranquillanti. Nello stesso momento il buspiron e altro blockers di 5-HT1A-retseptorov hanno l'effetto anxiolytic abbastanza efficace su altri modelli di frustrazione inquietante (Barrett e Vanover, 1993). I lavori recenti su topi con i geni inactivated di 5-NT1L-retseptorov anche testimoniano per vantaggio di un ruolo di questi recettori in pathogenesis di disagio e, forse, le depressioni (I parchi e al., 1998; Ramboz e al., 1998). D'altra parte, stimulators di alcuni i recettori serotonin, compreso 5-T2A--, disagio di causa di 5-HT2C-and 5-HT3-retseptorov in animali sia sperimentali, sia nella persona. Il ruolo di questi recettori e in modelli sperimentali di una depressione è supposto (per esempio, l'impotenza formata).
La persona di dati diretti su un ruolo serotonin in un pathogenesis di una depressione non ha ancora abbastanza. Nello stesso momento ci sono fatti clinici molto lucenti. Dunque, a pazienti con effetti di depressione di inibitori del ritorno serotonin la ricomprensione rapidamente sono eliminati alle influenze che calano serotonin il livello in un cervello. Siccome tali influenze servono, per esempio, il ricevimento di parachlorphenylalanine o le bevande che non contengono un tryptophan, ma ricco con amminoacidi neutrali (Delgado etal., 1990). È interessante che queste influenze in sé non provocano e non aggravano una depressione. Significa che il livello sufficiente di un serotonin in un cervello è necessario prima di tutto per efficienza di inibitori della sua cattura di ritorno.
Agenti che influenzano serotonin livello in tessuti
Negli esperimenti mandati a chiarificazione di un ruolo fisiologico di un serotonin è possibile applicare blockers di recettori serotonin o gli agenti che influenzano serotonin il livello in tessuti. Fino a recentemente nella maggioranza di lavori solo i farmaci del secondo dei gruppi menzionati sono stati meccanismi usati di azione di blockers i recettori serotonin sono stati studiati male.
All'atto della manutenzione bassa di un tryptophan in una dieta serotonin il livello in un cervello diminuisce, al contenuto alto di aumenti di tryptophan. Siccome la reazione di limitazione di sintesi di un serotonin è catalizzata, gli inibitori di questo enzima causano la caduta affilata di livello di un serotonin. Il più spesso usi un inibitore selettivo irreversibile di un triptofangidroksilaza parachlorphenylalanine. Sotto l'influenza di questa sostanza serotonin il livello considerevolmente e per molto tempo diminuisce, e la manutenzione di catecholamins non cambia.
Parachloramphetamine e altre anfetammine halogenated causano l'emissione di serotonin da thrombocytes e neuroni. In un cervello dopo che là viene la depressione lunga di riserve di serotonin. Le anfetammine di Halogenated sono largamente usate in lavori sperimentali. Due di loro fenfluramin e decsfenfluramin furono applicati come anorecsant, comunque in connessione con relazioni sulla loro azione tossica cardio nel 1998 loro negli Stati Uniti furono ritirati da vendita. Le conseguenze di uso di questi agenti sono fino alla fine non studiata. Nei neuroni cerebrali serotonin causano espresso e lungo (fino a parecchie settimane) la depressione di livello di un serotonin e il contenuto di proteine, specifiche per questi neuroni che un corriere di un serotonin e triptofangidroksilaza nello stesso momento diminuisce. Può dimostrare l'azione neurotoxin, comunque i segni di morte di neuroni sotto l'influenza di anfetammine halogenated non sono trovati. Derivati di triptamin con deputati supplementari in Indo-lino un anello (per esempio, 5.7-digidroksitriptamin; vedi il fico. 11.1) indubbiamente causano la morte i neuroni serotonin. L'introduzione di un 5.7-digidroksitriptamin un animale adulto è condotto a distruzione selettiva da fini serotonin, comunque i corpi di neuroni non soffre, e durante tempo le fini si rigenerano. Anzi, ad animali neonati sia le fini sia i corpi di neuroni serotonin e perciò il neogenesis non avviene periscono.
Gli inibitori del ritorno serotonin la ricomprensione, per esempio fluoxetine appartengono agli agenti che precisamente influenzano il trasferimento di serotonin. Il meccanismo della loro azione consiste in prolungamento di effetti del serotonin assegnato all'atto d'iniziazione delle fini nervose. Se insieme con questi farmaci per entrare in 5 hydroxytryptophan, allora le influenze di serotonin bruscamente amplificano. Gli inibitori del ritorno serotonin la ricomprensione sono uno degli antidepressivi più moderni e molto diffusi. L'inibitore del ritorno serotonin la ricomprensione, Noradrenalinum e Dofaminum sibutramin è applicato come anoreksant. In un organismo due metabolites attivi che, probabilmente, e hanno l'effetto terapeutico gli sono formati. Non è chiaro già, l'influenza su quale mediatore ha causato l'effetto di sibutramin.
Gli inibitori di MAO e Reserpinum appartengono a non gli agenti selettivi che influenzano serotonin il livello in tessuti. Gli inibitori di MAO bloccano la strada principale di un metabolismo di un serotonin, e Reserpinum causa la sua emissione da magazzino di neurone con il logoramento successivo. Tutti questi agenti conducono alla depressione espressa di manutenzione di serotonin, comunque nello stesso momento nello stesso grado anche il livello di diminuzioni di catecholamine. Perciò siccome gli agenti per gli inibitori di analisi farmacologici di MAO e Reserpinum sono usati di rado. Sono stati applicati in psichiatria: Reserpinum come narcoleptic, e inibitori MAO come antidepressivi.